

")
A propos
Construire un terrain de partage et de discussion autour des secrets de l’organe le plus complexe et mystérieux du vivant : tel est le but de ce blog dédié au cerveau. Des chercheurs en neurosciences y décryptent les avancées les plus importantes et prodigieuses, et vous emmènent à la découverte du système nerveux, de ses fonctions et de ses mystères. Lire ici l'éditorial du blog.
Contact : Giuseppe Gangarossa, giuseppe.gangarossa@univ-paris-diderot.fr
Twitter : @PeppeGanga
Les auteurs du blog

A la une

Comment prendre une décision rapide et adaptée dans un environnement complexe et changeant ? C’est une question à laquelle nous sommes tous confrontés, pris dans les flots d’informations parfois contradictoires propres à la vie moderne. De façon plus pragmatique, c’est un enjeu de survie pour tous les organismes vivants qui naviguent entre la nécessité de quérir des ressources et celle d’éviter les prédateurs. Dès lors, il semble raisonnable de postuler que l’évolution a favorisé le développement d’une architecture cérébrale permettant une prise de décision rapide et adaptée. En identifier les principes fonctionnels est un enjeu majeur des neurosciences contemporaines.

Comment choisir le bon chemin ? Une mission pour le cortex ? Jens Lelie/Unsplash, CC BY-SA
Classiquement, il a été considéré que les fonctions cognitives les plus avancées comme la capacité à raisonner, à planifier ou encore à élaborer des stratégies en vue de résoudre un problème devaient nécessairement s’incarner dans les régions du cerveau les plus évoluées.
De ce raisonnement découle un large intérêt des neuroscientifiques pour le cortex, et plus particulièrement pour sa partie dite préfrontale qui en constitue en effet la région la plus évoluée. Cet engouement, s’il est compréhensible, a néanmoins pour inconvénient d’occulter l’apport fonctionnel d’autres régions cérébrales.
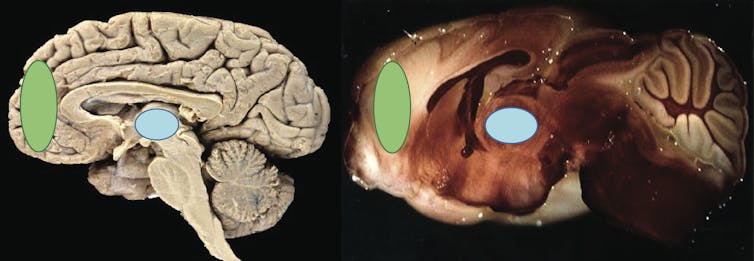 Chez l’homme (gauche) comme le rongeur (droite) le cortex préfrontal (vert) est la partie la plus antérieure du cerveau. Chez ces deux espèces, le thalamus (bleu) correspond à une région profonde, dite sous-corticale. Voir le paragraphe suivant pour cette dernière région.
Chez l’homme (gauche) comme le rongeur (droite) le cortex préfrontal (vert) est la partie la plus antérieure du cerveau. Chez ces deux espèces, le thalamus (bleu) correspond à une région profonde, dite sous-corticale. Voir le paragraphe suivant pour cette dernière région.
Deux constats permettent d’appréhender l’importance de cet apport. D’abord, des organismes au cortex plus rudimentaire que les primates sont également capables de prendre des décisions rapides et adaptées dont dépend leur survie.
Ensuite, le cortex préfrontal, comme les autres régions corticales, fait l’objet de projections anatomiques importantes de la part d’autres régions cérébrales, enfouies sous le cortex et apparues plus tôt dans l’évolution. Ces projections sont certainement appelées à jouer un rôle fonctionnel important.
Cortex préfrontal et « relais » thalamique ?
Une région sous-corticale particulière retient l’attention (ce n’est certes pas la seule !), il s’agit du thalamus qui est la source principale de l’innervation de l’ensemble du cortex, et particulièrement de sa subdivision préfrontale. Les liens anatomiques entre cortex et thalamus sont si étroits qu’à l’origine, les différentes régions corticales sont définies selon l’origine thalamique principale des connexions qui les innervent.
Pour comprendre cela, il est nécessaire de concevoir le thalamus comme une structure fortement différenciée qui est constituée d’une multitude de « noyaux » (soit des groupements de neurones) ayant chacun une connectivité corticale qui lui est propre. Ainsi, la partie dorsale du thalamus est la source majeure des afférences du cortex préfrontal tandis que d’autres noyaux thalamiques innervent le reste du cortex.
Initialement, le rôle du thalamus est conçu comme étant simplement celui d’un transfert passif de l’information vers l’étage cortical supérieur, où les opérations mentales complexes sont censées opérer. Selon cette vision classique, qui sera celle exposée dans les manuels pendant des décennies (ou dans l’article Wikimédia cité plus haut…), le rôle de cette structure ancienne s’apparente donc à celui d’un simple relais, qui n’apporte pas de contribution fonctionnelle originale.
De façon paradoxale, des faits cliniques et expérimentaux majeurs indiquent pourtant précocement que cette conception est manifestement incomplète, voire erronée, mais cela ne suffira pas à capter l’attention de la majorité des neuroscientifiques, plus préoccupés par les structures cérébrales dont le rôle fonctionnel apparaît plus immédiatement (par exemple, à ce jour près de 25 000 études sont publiées concernant le rôle du cortex dans les fonctions cognitives. Pourtant, dès le début des années 1980, le rôle du thalamus dans la mémoire est reconnu et quelques années plus tard, certaines conceptions théoriques issues de la neurophysiologie font valoir le fait que le cortex envoie en retour lui aussi des projections vers le thalamus et que donc cette dernière région ne saurait être limitée à un rôle de relais (les travaux et conceptions princeps de Guillery et Sherman, dont une perspective est désormais disponible).
Boucles thalamocorticales
Ces considérations finissant par trouver un certain écho, des études plus nombreuses voient le jour permettant d’établir que des interactions fonctionnelles sont à l’œuvre entre cortex et thalamus. Ainsi s’amorce un véritable changement de paradigme, puisque l’on passe progressivement d’une vision où le thalamus relaye les informations importantes au cortex pour un traitement approprié à celle où deux partenaires coopèrent, pour réaliser des opérations cognitives importantes.
Il faut néanmoins attendre les toutes dernières années et la disponibilité de nouvelles techniques à disposition des neurobiologistes pour véritablement s’attaquer aux principes fonctionnels qui sont à l’œuvre dans les « boucles » qui sont formées par les projections réciproques qui unissent cortex et thalamus.
 Les neurones du cortex préfrontal qui innervent le thalamus apparaissent en rouge. Seuls ces neurones corticaux expriment donc le récepteur muté qui permettra ensuite de les inhiber en injectant une substance inerte à l’animal. Ainsi nous pouvons examiner l’impact de l’inhibition de la voie corticothalamique sur la capacité à prendre une décision. De façon similaire, les neurones du thalamus qui innervent le cortex préfrontal peuvent également être inhibés.
Les neurones du cortex préfrontal qui innervent le thalamus apparaissent en rouge. Seuls ces neurones corticaux expriment donc le récepteur muté qui permettra ensuite de les inhiber en injectant une substance inerte à l’animal. Ainsi nous pouvons examiner l’impact de l’inhibition de la voie corticothalamique sur la capacité à prendre une décision. De façon similaire, les neurones du thalamus qui innervent le cortex préfrontal peuvent également être inhibés.
Ces techniques sont basées sur les progrès de la génétique et de l’ingénierie virale et permettent de prendre le contrôle de certains neurones d’intérêt que le chercheur pourra soit inhiber, soit au contraire activer de façon très sélective. Les techniques les plus avancées permettent de modifier des neurones d’intérêt en les dotant de récepteurs sensibles à certaines longueurs d’onde lumineuse (optogénétique) ou à des substances pharmacologiques exogènes à l’organisme (pharmacogénétique).
Mais l’intérêt principal de ces techniques est de permettre d’exercer un contrôle sur une catégorie bien définie de neurones avec une sélectivité sans précédent. En utilisant des vecteurs viraux aux propriétés complémentaires, on va en effet pouvoir cibler exclusivement les neurones thalamiques qui innervent le cortex ou bien au contraire uniquement les neurones corticaux qui innervent le thalamus. En bref, nous pouvons désormais étudier de façon spécifique les éléments constituants de ces fameuses boucles : les voies thalamocorticales (du thalamus vers le cortex) et les voies corticothalamiques (du cortex vers le thalamus).
Le choix du rat de laboratoire
Avec ces outils, nous pouvons donc désormais manipuler ces boucles thalamocorticales, mais une décision « rapide et adaptée » chez le rat de laboratoire, ça ressemble à quoi ?
Quand un mammifère prend une décision, puis effectue une action pour atteindre un but, il forme une représentation mentale de la valeur de la récompense escomptée et aussi du lien causal entre son action et l’effet qui en résulte. Quand ces deux critères sont établis, on parle d’un « comportement dirigé vers un but ». Ces capacités de représentation peuvent être étudiées au laboratoire par des procédures issues de la psychologie expérimentale.
Le principe général repose sur un apprentissage dit « instrumental » : l’animal apprend que deux actions différentes (pousser un levier ou bien une tige métallique) permettent l’obtention de deux récompenses alimentaires distinctes qui leur sont spécifiquement associées. Il apprend également le lien causal entre le fait de réaliser l’action et ce qui en découle. Des manipulations spécifiques vont nous permettre d’étudier comment l’animal peut mettre à jour sa représentation de ces deux attributs, valeur de la récompense et lien causal entre l’action et son effet.
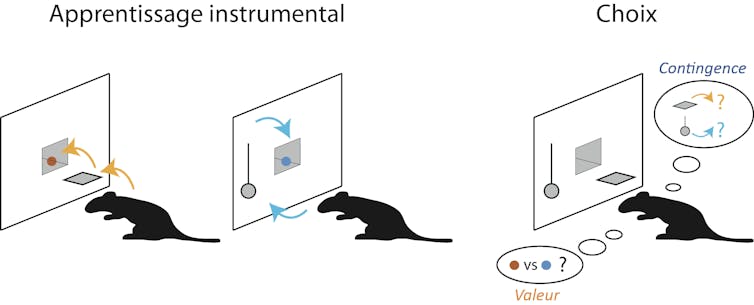 Pendant l’apprentissage instrumental, l’animal apprend que deux actions différentes (pousser un levier ou actionner une tige métallique) permettent d’obtenir deux récompenses spécifiques qui sont initialement également attractives pour l’animal. À la suite de manipulations spécifiques, l’animal peut être placé en situation de choix entre ces deux actions. Il devra fonder ce choix soit sur la valeur courante des récompenses (« maintenant que j’ai consommé beaucoup de pastilles marron, je préférerais obtenir la pastille bleue ») soit sur le lien causal, soit la contingence, entre l’action et son effet (« que se passe-t-il quand je presse le levier ou que j’actionne la tige métallique ? »).
Pendant l’apprentissage instrumental, l’animal apprend que deux actions différentes (pousser un levier ou actionner une tige métallique) permettent d’obtenir deux récompenses spécifiques qui sont initialement également attractives pour l’animal. À la suite de manipulations spécifiques, l’animal peut être placé en situation de choix entre ces deux actions. Il devra fonder ce choix soit sur la valeur courante des récompenses (« maintenant que j’ai consommé beaucoup de pastilles marron, je préférerais obtenir la pastille bleue ») soit sur le lien causal, soit la contingence, entre l’action et son effet (« que se passe-t-il quand je presse le levier ou que j’actionne la tige métallique ? »).
Dans le premier cas, on va changer temporairement la valeur de l’une des récompenses en permettant à l’animal de la consommer à volonté. Lorsqu’après cette phase, l’animal sera immédiatement placé en situation de faire un choix entre les deux actions, il va préférentiellement opter pour celle qui conduit à la récompense qui n’a pas été largement consommée au préalable. En effet l’appétence relative de la récompense largement consommée a diminué (si je viens de consommer un paquet de caramels mous, je préférerais probablement l’alternative d’une autre sucrerie si on me donne le choix entre ces deux options). Ensuite, on peut modifier l’apprentissage instrumental initial de façon subtile : presser le levier conduit toujours à la récompense qui lui a été précédemment associée mais presser la tige métallique n’est en revanche plus nécessaire car les récompenses qui lui étaient spécifiquement associées sont distribuées même si l’animal n’effectue aucune action. Placé à nouveau en situation de choix, l’animal va cette fois préférentiellement opter pour l’action qui permet effectivement d’obtenir la récompense plutôt que celle dont la relation de contingence avec la récompense a été ainsi dégradée.
Un rôle différent pour les voies thalamocorticales et corticothalamiques
Munis de l’ensemble de ces outils viraux et comportementaux, nous avons pu étudier au laboratoire le rôle fonctionnel des voies thalamocorticales et corticothalamiques de façon systématique dans la capacité à prendre une décision. Pour ce faire nous avons réalisé deux expériences complémentaires visant à inhiber de façon transitoire et spécifique par la méthode pharmacogénétique déjà évoquée les neurones thalamiques innervant le cortex préfrontal ou bien au contraire (chez d’autres rats) les neurones corticaux innervant le thalamus. À chaque fois, nous avons examiné le choix des animaux lorsque nous évaluons la valeur courante de la récompense, ou le lien causal entre l’action et son effet, soit les deux attributs principaux de la prise de décision.
Les résultats que nous avons obtenus permettent d’établir que ces voies ont un rôle complémentaire mais néanmoins différent : inhiber la voie corticothalamique empêche les animaux de fonder leur choix sur la valeur courante de la récompense, mais pas sur l’effet escompté de l’action. Inhiber la voie thalamocorticale est à l’inverse essentiellement délétère pour baser le choix sur le lien causal entre l’action et son effet.
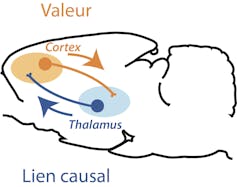 Schéma d’un cerveau de rat montrant que les deux attributs principaux de la prise de décision (valeur de l’action et contingence) sont implémentés de façons différentielles par les voies thalamocorticales et corticothalamiques.
Schéma d’un cerveau de rat montrant que les deux attributs principaux de la prise de décision (valeur de l’action et contingence) sont implémentés de façons différentielles par les voies thalamocorticales et corticothalamiques.
La direction des informations dans les circuits neuronaux
Ces résultats vont clairement à l’encontre de la vision réductionniste classique du thalamus conçu comme un relais puisqu’ils font au contraire apparaître le cortex préfrontal et le thalamus comme deux partenaires qui ont chacun une partition à tenir pour une prise de décision adaptée. De façon importante la direction de propagation de l’information apparaît dès lors comme une caractéristique fonctionnelle importante des circuits thalamocorticaux.
Cette caractéristique ouvre de nouvelles perspectives pour mieux appréhender les principes de fonctionnement des circuits neuronaux. En effet, de nombreuses régions cérébrales sont réciproquement connectées, et éprouver la généralité du principe de direction des informations apparaît comme un objectif important.
![]() Au-delà, l’enjeu reste de mieux comprendre le fonctionnement de notre cerveau et la disponibilité d’outils viraux appropriés permet désormais de questionner véritablement sa connectivité fonctionnelle. C’est une condition nécessaire pour mieux appréhender les troubles mentaux justement conçus comme des désordres de connectivité fonctionnelle entre aires cérébrales, comme c’est le cas de la schizophrénie par exemple.
Au-delà, l’enjeu reste de mieux comprendre le fonctionnement de notre cerveau et la disponibilité d’outils viraux appropriés permet désormais de questionner véritablement sa connectivité fonctionnelle. C’est une condition nécessaire pour mieux appréhender les troubles mentaux justement conçus comme des désordres de connectivité fonctionnelle entre aires cérébrales, comme c’est le cas de la schizophrénie par exemple.
Mathieu Wolff, chargé de recherches CNRS, université de Bordeaux
La version originale de cet article a été publiée sur The Conversation.
--------------------------------------------------------------------------------------------------------------------------------
Mathieu Wolff est chargé de recherches au CNRS. Depuis une douzaine d'années, il s'intéresse au rôle du thalamus dans les fonctions cognitives, une lubie acquise durant ses trois années de post-doctorat à l'université de Canterbury, à Christchurch (Nouvelle-Zélande). Actuellement, Mathieu travaille à Bordeaux dans l'équipe « Décision et Adaptation » de l'Institut de Neurosciences Cognitives et Intégratives d'Aquitaine (INCIA). Avec ses collègues, il s'intéresse aux interactions fonctionnelles entre le thalamus et différentes aires corticales en poursuivant l'idée que l'architecture thalamocorticale permet d'implémenter de façon idéale des processus cognitifs à la base des comportements adaptatifs. Pour ce faire, il développe des approches de neuroanatomie descriptive et fonctionnelle chez le rat qui sont couplées aux approches comportementales héritées de la psychologie expérimentale.